“Attention, attention, attention!”
—Zen Master Ikku’s answers when asked for the source of the highest wisdom
It helps to conceptualize meditation as an attentive art, so let’s start with meditation’s two basic categories. The first kind employs an effortful, sustained attention. This variety of concentrative meditation is the easiest to understand. It’s what we begin with and what we return to frequently during meditation. Concentration implies that we narrow our focus voluntarily. We choose this target and exclude others. When we intensify concentration in such a “top-down” executive manner, it can later evolve into the absorptions, the jhānas. When we describe this focused, top-down form of meditation as implicitly Self-referential, it means only that we ourselves are doing it. We’re the active agents in charge. Even when we’re following our breath down to its slightest movements in the lower abdomen, our Self is still in there “paying attention” in this willful, voluntary manner.
(We’ll use a capital S to suggest that this personal Self creates many large problems.)
The second category, receptive meditation, is less obvious and less easily defined. It begins as a bare, choiceless awareness. Why is this awareness called choiceless? Because no person is in there choosing what to focus on. Therefore, this kind of involuntary meditation is more effortless. Its modes of “bottom-up” attentive processing are wide open to include any stimulus that might come unexpectedly from anywhere. A receptive awareness so universal in scope is basically other-referential. Subsequent sections expand on several important implications of such “bottom-up” receptivities. Increasingly, they are poised to shift into intuitive modes of comprehension—into insights of various depths (hence, its common name in Western Buddhism: Insight Meditation, from the Pali vipassanā).
Our two ways of perceiving reality
Two processing streams are involved when we perceive reality. The first is our familiar egocentric processing system. Take, as an example, someone who looks down toward a table to reach for a nearby apple. Photons from that apple are transformed by the eyes into impulses coursing toward the back of that person’s brain. In the opening milliseconds, the brain represents this apple’s raw visual image in 3-dimensional spatial coordinates. This apple-message is referred directly back to circuits representing his or her own physical Self. This somatic Self remains the central axis receiving all such Self-referent 3-D perceptions. This is an automatically Self-centered, egocentric reconstruction of a real apple. It’s also the obvious way that we’re each continually reminded: I am this unique someone who is looking at that thing down there.
But there’s a second, covert way of perceiving reality; we’re normally totally unaware of it. Its mode of perception is other-centered, not Self-centered. Therefore, most research reports in psychology now refer to it as allocentric perception, from the Greek, allos, “other.” Allocentric accurately describes the seemingly objective, externalized perspective that first creates images of objects in the outside world. The apples it sees co-exist in relation to other apples “out there,” not with reference back to our body in the center.
It’s almost as if the allocentric functions exist on some other planet, having no need for us back on earth to be witnessing it.
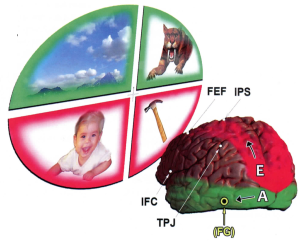
Functional anatomy of two streams, E & A
Let’s now return to take up the first, egocentric processing stream in greater detail. [See figure, below.] Here, your vantage point is from a position behind the brain’s left hemisphere. This brain is shown gazing up and off to the left into quadrants of outside scenery. The egocentric processing pathway (E) is shown pursuing its upward trajectory. Along the way, it overlaps with the two major modules for our top-down visual attention. These are the intraparietal sulcus (IPS) and the frontal eye field (FEF). On this upper trajectory, it’s clear that our Self-centered stream flows first through the parietal lobe. Note what this implies: our top-down form of attentive processing is inherently linked with that of our own body, our soma, our personal physical self. Up here, in the parietal lobe, this Self-referential stream enlists the services of two highly personal special senses. One is proprioception—our subliminal perception of exactly where and how our body parts exist in space. The second is our sense of touch. Note that we use proprioception and touch to help us handle things close to our body.
In a top-down proprioceptive and tactile decision, the functions are not only intimately Self-centered (egocentric), their metrics are also action-oriented. Indeed, when objects are positioned down here close in front of us, we’re often looking down at them below the horizon of our gaze, and our hands are working on them with tools. How do we localize the position of a tangible object? We refer it back to the axis of our “body image,” a schema divided into left and right halves, for example.
In contrast, the allocentric stream (A), pursues a trajectory lower down. This processing stream flows through networks down in the temporal lobe. As soon as they register an object farther away, they identify it, aided by their pattern recognition functions. Moreover, this lower processing stream is also poised to infuse meanings and values into whatever it perceives out there. It’s hard for us to get our mind around the notion of this allocentric stream because it begins so anonymously. It’s almost as though its functions exist on some other planet, having no need for us back on earth to be witnessing it. Its mode also seems inherently objective. Why? Because none of our private subjectivities are involved in it. In this respect, note that it enlists the vital associations of two other special senses—vision and hearing. These are the two special senses we use when we wish to detect objects and evaluate them at much greater distances from our bodies.
Note how this lower stream flows downward first, then runs through the temporal cortex on its way toward the lower frontal regions. Along the way, its trajectory is accessible to the two other modules of attention that serve our bottom-up attentive functions: the temporo-parietal junction (TPJ) and the inferior frontal cortex (IFC). Keep in mind that these temporal lobe networks are designed for meaningful interpretations based on seeing and hearing.
Imagine that the right side of the brain struck a bargain eons ago with its opposite hemisphere: you handle language, but I’m going to take our subtler kinds of attention.
The “crossings” of functions
By now, the general public tends to be aware that the left hemisphere plays the dominant role in the way people speak, read, and listen to language. This left hemisphere also handles fine details more efficiently, using its fine-grain discrimination functions. Moreover, when the left hemisphere looks out, it tends to see things that lie over in the opposite right side of the environment.
Why do so many of our functions seem to cross over to—and return from—the opposite side? Brain pathways often do cross over when they transmit messages entering from the opposite side of the outside world or the opposite side of our body. Indeed similar crossings also point to the way that our temporo-occipital region (low down in the back of the brain) attends most efficiently to visual stimuli that enter our upper visual fields. Referring again to the figure, this explains why the green allocentric stream (A), which flows through the lower regions of the brain, is shown receiving visual information from the green-framed scenery in the upper visual quadrants. In contrast, the upper red egocentric stream (E) receives visual information from nearby targets framed by red in the lower visual fields.
However, there’s another important point: the lower pathways in the right hemisphere play the predominant role in certain key aspects of our attention. This information is crucial for meditators to be aware of. Because when you want to further refine both your involuntary forms of bottom-up attention and the interpretive nuances of their nearby allocentric processing, you’ll be chiefly exercising these automatic intuitive functions of your right hemisphere.
Moreover, this right hemisphere takes on another responsibility. It attends not only to the opposite (left) visual field but also to the right side of the outside environment. Imagine that the right side had struck the following kind of bargain eons ago with its opposite hemisphere: OK, you can handle language over on your left side, but I’m going to take credit for our subtler kinds of attention. These are the skills poised to service unexpected needs to attend to both sides of the environment. Today, what is the result of such a pre-emptive decision? A right hemisphere that enables us to attend—in an other-referential manner—to the outside world on both sides, simultaneously. Consider how this sets the stage for higher states of attention to develop.
Note also the survival advantage. Hearing detects some rustling noises. Vision glimpses the potential predator. Such a brain can identify a fearsome beast while it still remains a relatively long distance away from one’s body.
Why is that saber-tooth tiger pictured in the upper field of vision? To identify the beast instantly, one needs to discriminate the tiger’s color patterns from those of the adjacent green foliage. Nerve cells specialized for color discrimination are located in the fusiform gyrus (FG) at an early stage along the allocentric pathway. From there, the pathway diverges into the pattern recognition regions of the rest of the temporal lobe. Later, a grateful survivor might look up above the horizontal and off to the left, to enjoy the mountain scenery and distant clouds.
Perhaps you’re also wondering why a baby and a hammer are shown nearby in the lower visual fields. Everyone worries about the possibility of actually dropping a baby. It takes top-down decisions to pick up a baby and balance it on your lap. You pay careful executive attention to how you position your hands. It also takes a lot of skill to aim a hammer when you hold a small nail. That hammer head must come down smack on the nail head, or your fingers are history.
In summary, thus far, evolution seems to have wired our circuits in the following manner:
- The lower right hemisphere plays a dominant role in our bottom-up, reflexive attentive processing. In this respect, its functions are reminiscent of the way corresponding regions over on the left side were designed to exercise its dominant role in our motor speech and comprehension of language.
- The upper, parietal pathway in each hemisphere is designed to service our Self-centered behaviors, especially when we look down to grasp and manipulate things held close to our body.
- In contrast, the temporal lobe pathways lower down are tuned to attend most efficiently to other messages entering from the more distant environment. Visually, the temporal lobe is more sensitive to stimuli that can enter from the scenery higher up above our horizon of gaze. Moreover, it is poised to interpret sound stimuli that arise from anywhere.
The l-Me-Mine Triad
Our “I” is arrogant and aggressive. Our “Me” is its vulnerable partner, trembling with fear.
It’s useful to envision three main interactive components of our personal Self, in order to understand why they go on to have maladaptive consequences.
The I-Me-Mine triad operates within certain overlapping premises. The I itself has a tangible core. Our soma obviously exists physically, and it obviously feels emotions of all kinds. Moreover, the I knows that it is aware, knows that it acts, thinks, and personifies various roles.
Things happen to the Me. Things hurt Me, both my soma and psyche. This Me is the target for every sling and arrow of outrageous fortune.
Finally, the Mine represents our greedy, possessive self. All private thoughts, opinions, and body parts are mine. Don’t invade My turf. Remember: these prized possessions are Mine.
This complex triad clearly serves useful and adaptive ends. But many maladaptive dysfunctions are associated with such a Self. For example, our I is arrogant and aggressive. Our Me is its vulnerable partner, trembling with fear. It feels besieged and gets battered. Finally, our Mine is easily captured. It covets and craves material goods. It clutches at other people. An especially pernicious habit is the way our Mine cherishes and clings to old biased opinions. These are the kinds of “a-b-c’s” that suggest the everyday liabilities of our I-Me-Mine.
Our active “resting” brain
Only recently did neuroimaging researchers actually pay volunteers to do nothing. The subjects were instructed to relax yet stay awake. The resulting passive setting resembles a preamble to several of the more relaxed styles of meditation—but only in the sense that these research subjects were being encouraged to begin with few top-down goals in their conscious minds.
The subjects’ earlier PET scans and more recent fMRI scans (functional magnetic resonance imaging) show two big regions of activity, deep inside the brain, along the inner surfaces of its right and left halves. One big “hot spot” lies in the medial prefrontal cortex, again on each side. The other lies in the deep medial posterior parietal region, also on each side. A smaller “hot spot” lies on the outside of the brain in the lower parietal cortex on each side. It involves each angular gyrus, a crossroad in the egocentric pathway. Is this truly a “resting brain?” If so, these particular regions are very active.
Later, researchers asked their subjects to engage in some precisely Self-related activity (such as introspecting into their very own personality). Now, each large medial frontal hot spot became even more active than it was during the resting baseline period. Later still, the subjects could be asked to dig back into their own memory of a particular event, one in which they were personally involved. Now, when the subjects re-imaged the circumstantial details of that particular scene, their parietal regions also became even more active than they had been at rest.
We might briefly summarize this evidence about the already hot spots existing inside the so-called “resting” brain: 1. Some regions are further activated when that person engages in inturned, Self-relational functions; and 2. Others also become turned on more when such Self-inspired acts become recorded in the context of other circumstantial details drawn from the outside world.
These observations suggest that the brain has ways to enlist a spectrum of “self-othering” functions. What practical purpose might be served by blending the contributions of these Self-centered frontal networks with those supplied by their Self-other parietal partners? It’s possible to envision such interactions as the source for entries to our ongoing, daily journal. Each of us stores a lifetime of personalized events as we go about our ordinary, everyday activities. They take the form of “who-what-where-and-when” entries. They help us remember: “I graduated from this high school, and with these guys and girls, umpteen years ago. This church is where my wife and I were married. This is where we went on our honeymoon. This is how I reconstruct my life narrative when I explain who I am to a stranger. Each time I need to navigate within my surroundings, I can consult this vast storehouse of my detailed memories.”
Each time our attentive regions are enhanced, its shifts can deactivate our Self-referential regions.
These memory traces (engrams) record our ongoing private sense of Self. They need to be indexed, distributed and stored in ways that are readily accessible. We require baseline resources of metabolic energy to maintain these journal entries. We also need standby reserves of energy instantly to select and recall which precise entries match our most urgent needs.
Our “monkey mind”
We’ve jam-packed these networks with trivial entries accumulated over a lifetime. We also keep them preoccupied with dire worries about fictitious events that we continue to project into an imaginary future. Is it any wonder that thoughts intrude when we meditate? Where do such frantic fears, plans, and scenic details come from? They reflect the over-activities of distributed networks that stay ready to be engaged, even when we’d much prefer otherwise.
“Monkey-mind” describes these intrusive thoughts. They leap around between past, present, and future, jump from one branching topic to another. In contrast, the emphasis in Zen is on no-thought meditation, mushin. One ancient adage concludes: “Forget about the branches— get to the underlying root.”
Attention triggers deactivation
To get to the root means going deeper in the brain. The next research findings become more complicated, because sometimes researchers presented their resting subjects with a significant fresh task to perform. Now, those three previous hot spots became “cooler.” Those three active spots became deactivated. Let’s not be mislead by how much processing was obviously required to solve the whole task itself. Let’s focus first on just what’s needed in the earliest milliseconds: a task must first capture our attention in order to be considered significant. First, we need to attend to it. We also need to sustain this attentive interest to process the task appropriately.
The two-word phrase, attentive processing, condenses this practical principle. The order of these two words is instructive. Attention comes first. Attention is the vanguard. Each sharp point of attention is the salient event that impales its target. Only when attention helps focus on any new task can we then mobilize our subsequent processing. Indeed, as soon as our brain confronts a fresh demand to attend, the fMRI evidence can be interpreted to suggest that the brain’s former active hot spots react by becoming cooler (“deactivated.”) Such a shift could help cut through all the trivia, and point directly toward those few circuits that are essential to solve the immediate task at hand.
Brain’s slow, spontaneous intrinsic rhythms
Other intriguing things happen in the so-called “resting brain” when researchers stop assigning new tasks and simply observe a resting subject’s functional MRI scan for as long as five minutes. Sometimes the hot spots get a little hotter. At this same time, the different cool spots get a little cooler. After a while, the reverse occurs: when those different cool spots now get a little hotter, the previous hot spots now get a little cooler. A reciprocal “see-saw” effect keeps recurring. It turns out that one of these reciprocal effects increases the activity of those outer cortical regions involved in attentive functions; simultaneously it decreases the activity of those mostly medial fronto-parietal regions that we’ve seen are especially involved in a spectrum of Self-other functions.
These up-and-down fluctuations shift spontaneously back and forth between Self and attention three or four times a minute, on their own. Is such a slow, spontaneous rhythm of interest to meditators? Yes, because many Buddhist practices are dually oriented: 1. toward enhancing attentiveness; 2. toward decreasing the pejorative influences of the maladaptive Self. This recent fMRI evidence suggests that our normal brain has these same innate reciprocal capacities: each time our attentive regions are enhanced, its shifts can deactivate our Self- referential regions.
The amplitudes of attention, enhanced and sensitized by long meditative training, suddenly captured by an unexpected stimulus: the personal Self can drop out.
Note: we’re not referring to our usual brain waves, the kinds recorded in electroencephalograms (EEGs). EEG rhythms go up and down at much faster rates, multiple times each second. Instead, we’re observing an ultraslow fundamental rhythm of the human brain. This kind of metabolic-bioelectric rhythm fluctuates even more slowly than does our breathing rate of around eighteen times a minute. Its complex origins are still under study.
The implications of triggering events
Zen Master Ikku emphasized attention. When he was 26, what precipitated his sudden breakthrough into a non-dual experience of selfless insight? The auditory trigger for Ikku was the unexpected “caw” when a crow flew overhead. Zen annals record many similar episodes when a triggering stimulus abruptly captured attention and precipitated kensho-satori.
When did Siddhartha finally became enlightened? Legends tell us that it was when he looked up into the pre-dawn sky and saw the planet Venus. (Often overlooked is the way that six earlier years of rigorous spiritual training were the prelude to the Buddha’s supreme awakening by this “morning star.”) The triggering stimulus serves only as the “last straw” on an already overloaded (and sensitized) camel’s back.
We wonder: What makes an otherwise commonplace stimulus so effective, and only at a particular time? One plausible working hypothesis returns us to the brain’s reciprocal shifts between two of its modes: attentiveness and Self-centeredness. Let the amplitudes of attention become enhanced and sensitized by long meditative training, and then suddenly captured by an unexpected fresh stimulus. Now, finally, the personal Self can drop out of the mental field of consciousness.
That alone would not suffice. Simultaneously, attentive mechanisms need also to react selectively, in ways that liberate bottom-up attention as the vanguard of allocentric processing. Unleashed from their subordinate role, these lower networks can resonate in harmony with the whole outside environment. Allocentric perception (now heightened) can perceive its outside setting as a world both transformed and transfigured, as an environment manifestly “realer than real.” Finally, vision and hearing can register direct, selfless, impressions. They are interpreted as absolute perfection. No limbic-driven fears or other I-Me-Mine intrusions distort this timeless state.
Thalamic roots of selfhood
How could kensho-satori also dissolve every last personal trace of fear, perceive all things afresh, as THEY REALLY are? To answer, we need to ask why those normal hot spots in our cortex stay so highly active, even at rest, and understand which circuits drive all our monkey-mind wandering. Much of this usual internal dialogue is driven by deep connections with our limbic system. These limbic circuits keep transmitting their legacy of ancient instinctual angst in ways that over-condition our Self.
Yet, wouldn’t consciousness itself drop out if the Self and all its limbic overtones suddenly dissolved? No. Consciousness emerges transparently clear, fresh, stabilized. What does drop out in this state is the clutter referable to our Self-centered I-Me-Mine and its past accumulations of narrative trivia.
Let’s now pursue many of the questions raised above to their deeply rooted answers farther down in the dorsal thalamus.
- Three limbic nuclei reside in this upper tier of the thalamus. Not only do they connect with the limbic system, their circuits also transmit its affective biases up to drive the larger, Self-referential cortical hot spots. Extraordinary things can occur when certain sequences of events all happen to fall into place at just the right moment.
- The lateral posterior nucleus is another key nucleus in this dorsal thalamus. It connects intimately with the cortex of the superior parietal lobule. This is our somatosensory association cortex. It helps create the schema that integrates separate parts of our soma (our physical Self) into a functioning body image.
- The large dorsal pulvinar resides at the back end of the thalamus. Its vital associations contribute the normal sense of salience to our blend of Self-centered perceptions. It also interacts with both the large medial parietal and smaller angular gyrus hot spots of the parietal lobe.
What role could the ventral pulvinar and its projections play during kensho-satori as their messages continue to stream forward through networks in the temporal lobes? These pathways—now unleashed from inhibition—remain to sponsor the enhanced sense of values associated with insightful bottom-up, attentive allocentric processing.
GABA inhibition
How can a brain be so selective about what it activates and deactivates? The reticular nucleus of the thalamus is crucial in this regard. It embraces the whole thalamus, acting like a shield. It is composed of a thin gauze network of GABA nerve cells. (GABA stands for gamma aminobuteric acid.) GABA is our brain’s major inhibitory neurotransmitter. Its role is to stop over-active nerve cells from firing excessively. Each time the reticular nucleus detects too much activity in the circuits that normally oscillate between thalamus and cortex, it releases its GABA to check this imbalance.
These pages emphasize the normal excitatory and inhibitory interactions between the limbic system, thalamus, and cortex. Their oscillations are not something esoteric just for mystics to tap into. Similar shifts go on each time you and I lie down at night and start dropping off into our normal slow wave sleep. At this time, as the reticular nucleus fires, its GABA shuts down the sensory transmissions relating to Self, and our monkey- mind finally stops.
In closing
This working hypothesis suggests how the meditative training of attention influences events down at the thalamic level that set the stage for the sudden breakthrough into the selfless insight of kensho. The resulting deactivations briefly dissolve both the egocentric overconditionings of the psychic Self and their links referable to the body schema of the physical Self. Liberated simultaneously, and selectively, are many functions of the allocentric pathway that stream forward through the temporal lobe.
Rarely, after the long and regular practice of training attention, you too might have occasion to look up and glimpse a bright, distant star, or perhaps to hear the penetrating sound of a bird call. Extraordinary things can occur when certain sequences of events all happen to fall into place at just the right moment. Yet, all too often, wandering thoughts and unbidden images clutter our minds. Covert influences from an overconditioned limbic system distract us with all their excess baggage.
A useful old Chinese Zen phrase is wu shin. It points toward our becoming empty of all such counterproductive interactions between heart and mind. It means neither being swept away by excess limbic baggage nor captured by cognitive concepts.
Does contemporary Zen include a prescription comparable to wu shin? If so, perhaps it might begin something like this: Just sit quietly. Be very patient. Learn how to let go. Allow your disquietudes to drop off. Open up into your receptive and inherently intuitive modes of meditation. Observe what happens. Remain so open that when thoughts come in the front door, they soon leave out the back door. Following Master Shunryu Suzuki’s advice, “Just don’t serve them tea.”
James H. Austin, M.D., studied Zen in Kyoto, Japan with Kobori-Roshi while pursuing a career as an academic neurologist. This article is condensed from Dr. Austin’s course at BCBS on May 2, 2009 and from his book Selfless Insight: Zen and the Meditative Transformations of Consciousness.